Санкт-Петербург, 2002 г.
РАЗДЕЛ 2. ТОКСИКОДИНАМИКА
ГЛАВА 2.2.
Механизмы цитотоксичности
В основе токсического действия веществ лежит повреждение клеток, сопровождающееся их функциональными, либо структурно-функциональными изменения. Разнообразие формирующихся при этом эффектов со стороны целостного организма обусловлено сложностью организации клеток, многообразием клеточных форм, составляющих организм. Сформировавшиеся в процессе эволюции особенности структуры и функции отдельных клеточных типов, формирующих различные органы и ткани, настолько существенны, что чувствительность различных клеток к токсикантам может отличаться в тысячи раз. Тем не менее, живое объединено общностью фундаментальных свойств (см. выше), а это позволяет выделить и некоторые общие механизмы, лежащие в основе цитотоксического действия ксенобиотиков. К числу важнейших можно отнести следующие:
- нарушение энергетического обмена;
- нарушение гомеостаза внутриклеточного кальция;
- активация свободно-радикальных процессов в клетке;
- нарушение процессов синтеза белка и клеточного деления;
- повреждение клеточных мембран;
Необходимо отметить, что все эти механизмы тесно связаны друг с другом. Порой один из них является пусковым, но в дальнейшем особую значимость для судьбы поврежденной клетки приобретают другие. Очень часто два или несколько из упомянутых механизмов связаны между собой по типу "порочного круга". В этой связи их выделение носит искусственный характер и оправдывается только целями данного издания.
1. Нарушение процессов биоэнергетики
1.1. Системы энергообеспечения клетки
Если между двумя веществами осуществляется взаимодействие, например:
А + В
то существует и некая константа равновесия, характеризующая силу этого взаимодействия:
К = [ АВ] /([ А] + [ В] ).
Если К = 1, то в равновесном состоянии компоненты реакции А (или В) и АВ находятся в смеси в одинаковых концентрациях. Если К > 1, то в смеси больше продукта реакции АВ, если К < 1, то, напротив, больше исходных ингредиентов А и В. Спонтанное взаимодействие А и В с образованием АВ происходит только тогда, когда К - велика.
Энергетика процесса взаимодействия веществ может быть охарактеризована следующим уравнением:
-RTlnKa =
Ка - константа равновесия;
R - газовая константа (1,987 кал/град/моль);
Т - абсолютная температура;
G - свободная энергия;
Н - энтальпия (энергосодержание веществ);
S - энтропия (степень неупорядоченности системы).
Спонтанно протекают реакции, при которых свободная энергия системы в процессе преобразования уменьшается. Это может быть следствием двух явлений: уменьшение энтальпии или увеличения энтропии системы. В первом случае говорят об экзотермической реакции (
Х + АТФ
Х-АДФ + У
В этом случае эндэргический процесс:
Х + У
становится возможным за счет сопряженного экзэргического процесса:
АТФ
в ходе которого образуется промежуточный активный продукт Х-АДФ.
Жизнь клеток и макроорганизмов есть постоянный процесс синтеза сложных молекул (нуклеиновых кислот, белков, полисахаридов, липидов и т.д.), то есть структур с достаточно высокой энтальпией и низкой энтропией. Образование таких молекул означает увеличения свободной энергии системы. Следовательно синтетические процессы невозможны без одновременного протекания экзэргических реакций, обеспечивающих высвобождение энергии, поступающей из окружающей среды и запасенной в форме химических соединений. Основным видом таких реакций в организме является гидролитическое расщепление богатых энергией веществ, содержащих пирофосфатные связи (макроэрги). К их числу относятся: аденозинтрифосфорная кислота (АТФ), аденозиндифосфорная кислота (АДФ), гуанозинтрифосфорная кислота (ГТФ_, цитозинтрифосфорная кислота (ЦТФ), уридинтрифосфорная кислота (УТФ), ацилфосфаты и др. В конечном итоге энергетические возможности клетки определяются запасами макроэргов и, в первую очередь, АТФ.
АТФ постоянно синтезируется в ходе двух клеточных процессов: гликолиза и биологического окисления субстратов (клеточное дыхание). Оба находятся в тесном взаимодействии. Гликолиз проходит в цитоплазме клеток. Дыхание, основной путь образования макроэргов, осуществляется в митохондриях. В процессе дыхания, благодаря совместному действию большого числа ферментов, соответствующие субстраты, образующиеся при метаболизме белков, жиров, углеводов, поступающих с пищей, окисляются кислородом, также поступающим из окружающей среды, с образованием СО2, Н2О и энергии, выделяющейся в форме тепла и запасаемой в молекулах АТФ. Процессы, обеспечивающие поддержания определенного уровня АТФ в клетках организма, составляют основу и сущность их энергетического обмена.
Наиболее уязвимыми для действия токсикантов элементами биологической системы, обеспечивающей образование макроэргов в организме, являются: механизмы биологического окисления (ферменты цикла трикарбоновых кислот, ферменты дыхательной цепи), механизмы сопряжение биологического окисления и фосфорилирования (образования АТФ из АДФ и фосфата), механизмы доставки кислорода к клеткам кровью (рисунок 1).
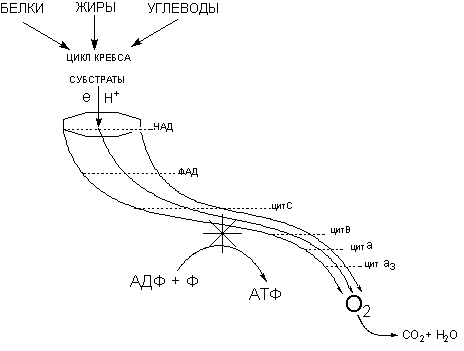
Рисунок 1. Модель процесса клеточного дыхания. Энергия движения электронов и протонов по цепи дыхательных ферментов обеспечивает работу механизма сопряжения процесса фосфорилирования (кумуляция энергии)
Угнетение активности энзимов гликолиза редко приводит к существенному нарушению энергетического обмена, так как метаболизм других веществ (жиров, белков) может компенсировать этот вид нарушений.
1.2. Механизмы токсического повреждения систем энергообеспечения клетки
Нарушение процессов биоэнергетики приводит к повреждению биологических систем, вплоть до их гибели. Токсиканты могут влиять на энергетические процессы различными способами. Некоторые примеры представлены на рисунке 2.
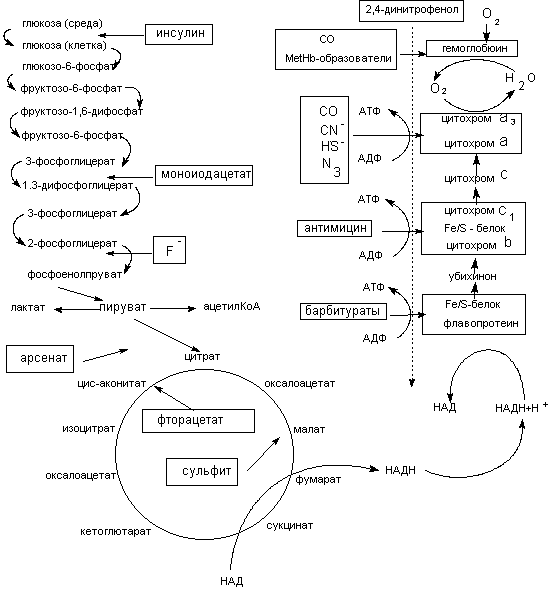
Рисунок 2. Механизмы действия токсикантов на биоэнергетические процессы
Мышьяк, ртуть, их органические и неорганические соединения, другие тяжелые металлы, иодацетат и проч., могут взаимодействовать с сульфгидрильными группами целого ряда энзимов гликолиза и цикла трикарбоновых кислот и подавлять их активность (сульфгидрильные яды). Достаточно избирательно действуют вещества-аналоги природных субстратов (фторацетат, оксид углерода и т.д.). Они взаимодействуют с активными центрами соответствующих энзимов, угнетая отдельные этапы процесса энергообеспечения клеток.
Ингибирование энзимов цикла трикарбоновых кислот (ТК) является пагубным для клетки. Так, некоторые фторированные спирты и фторкарбоновые кислоты (ингибиторы ферментов цикла ТК), являются чрезвычайно токсичными соединениями, вызывающими смертельное отравление в дозах нескольких миллиграмм на килограмм массы.
Большое токсикологическое значение имеют вещества, действующие на цепь дыхательных ферментов. Токсиканты, блокирующие элементы цепи дыхательных ферментов (цианиды, сульфиды), могут в течение нескольких минут привести организм к гибели. Первая ступень цепи дыхательных ферментов чувствительна к седативным средствам (барбитураты).
Некоторые вещества способны разобщать процессы биологического окисления и фосфорилирования. Такими свойствами обладают, как правило, липофильные соединения, содержащие фенольную группировку в молекуле и являющиеся слабыми органическими кислотами: 2,4-динитрофенол (ДНФ), динитро-о-крезол, хлорфенолы, дикумарол, а также салицилаты, арсенаты, тироксин и др. (рисунок 3). Самым активным разобщителем является 3,5-дитретбутил-4-гидроксибензилиден-малононитрил (SКF6847). Это вещество примерно в 1800 раз активнее 2,4-ДНФ и уже в концентрации 0,2 моль на 1 моль цитохрома вызывает максимальное разобщение.
in vitro "разобщители" стимулируют скорость потребления кислорода митохондриями и индуцируют активность АТФазы. in vivo в результате их действия значительно активируются процесс клеточного дыхания, однако содержание АТФ в клетках при этом снижается. Образующаяся в ходе окисления субстратов энергия рассеивается в форме тепла, температура тела отравленного резко повышается, учащается дыхание, появляются тошнота, обильное выделение пота, развивается острая сердечная недостаточность, кома. Интоксикация развивается бурно и заканчивается быстрой гибелью пострадавшего.
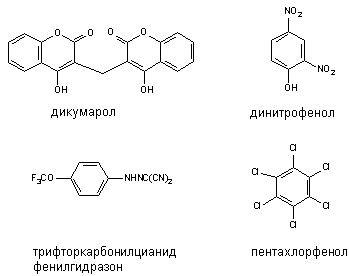
Рисунок 3. Структура некоторых разобщителей процесса окислительного фосфорилирования
Механизм действия разобщителей до конца не выяснен. Полагают, что они облегчают переход протонов непосредственно через мембрану из митохондрий в цитоплазму. Двигателем же процесса образования АТФ из АДФ и неорганического фосфата как раз и является градиент протонов по обе стороны мембраны митохондрий, не проницаемой для Н+, поддерживаемый реакциями биологического окисления.
Наконец еще одним механизмом нарушения энергообеспечения клетки является угнетение митохондриальной транслоказы, обеспечивающей транспорт ситезированной АТФ из митохондрий в цитоплазму. Таким образом, действует, в частности атрактилозид, вещество выделяемое из растения Distel Atractylis gummifera, произрастающего в Среднеземноморье (рисунок 4).
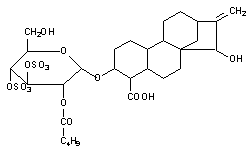
Рисунок 4. Структура атрактилозида
Непрямым механизмом цитотоксического действия является понижение токсикантами парциального давления кислорода в тканях.
Нехватку кислорода человек и другие млекопитающие могут переносить только несколько минут. Особенно чувствительными к аноксии являются клетки органов с интенсивным энергообменом (таблица 1).
Таблица 1. Потребление кислорода различными органами крысы
>Орган |
>Скорость потребления О2 (мл/г ткани мин) |
Скелетная мускулатура |
>0,875 |
Кожа |
>0,416 |
Кости |
>0,153 |
Кровь |
>0,025 |
Печень |
>2,010 |
Почки |
>1,840 |
Мозг |
>4,120 |
Легкие |
>1,250 |
Сердце |
>1,930 |
(Field et al., 1939)
Высоко чувствителен к нарушению процессов клеточного дыхания головной мозг. Если периферические ткани способны переживать (хотя и с нарушениями функций) частичную нехватку кислорода в течение нескольких часов, то необратимые изменения в ЦНС наступают спустя 4 - 5 минут после полного прекращения снабжения нейронов кислородом. Поэтому токсиканты, нарушающие кислородтранспортные функции крови также весьма токсичны (оксид углерода, нитро-, аминосоединения и пр.). Оксид углерода связывается с двухвалентным железом гема с образованием карбоксигемоглобина, метгемоглобионообразователи переводят железо гема из двухвалентной в трехвалентную форму. В обоих случаях гемоглобин утрачивает способность связывать кислород в легких и транспортировать его в ткани.
2. Нарушение гомеостаза внутриклеточного кальция
Роль кальция, как регулятора физиологических процессов, протекающих в клетке, хорошо известна. Нарушение внутриклеточного гомеостаза этого иона, сопровождающееся существенным повышением его концентрации в цитоплазме клетки, лежит, как полагают, в основе механизма клеточной гибели при целом ряде патологических состояний таких как ишемия, аутоиммунные процессы и т.д. Цитотоксическое действие самых разных токсикантов (цианидов, четыреххлористого углерода, свинца, метилртути, хлордекона, оловоорганических соединений, алкилирующих агентов, диоксина и т.д.) также полагают, хотя бы отчасти, связано с повышением уровня кальция внутри клеток.
В норме низкая концентрация кальция в цитоплазме поддерживается механизмами его активного транспорта через биомембраны и системой его компартментализации (накопление в клеточных органеллах) (рисунок 5).
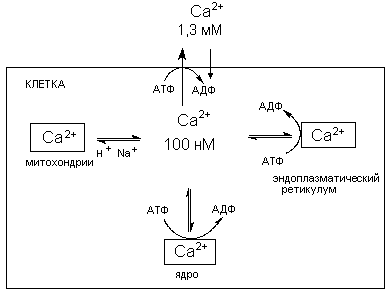
Рисунок 5. Схема регуляции содержания ионов кальция в цитоплазме.
Обычно поступление Са2+ в клетку по градиенту концентрации уравновешивается его активным выведением, осуществляемым при участии Са2+-транспортной АТФазы в печени и эритроцитах и Na+/Ca2+ обменом в возбудимых тканях. Кроме того, уровень кальция регулируется его секвенированием в клеточных органелдах (ядре, митохондриях, эндоплазматическом ретикулуме), а также связыванием внутриклеточными белками.
Нарушение механизмов поддержания гомеостаза внутриклеточного кальция, как это видно из представленной схемы обмена иона, может стать следствием:
- повреждения биологических мембран и усиления их проницаемости для ионов;
- нарушения биоэнергетики клетки, приводящие к истощению запасов макроэрогов;
- изменения функционального состояния белковых комплексов (в том числе путем действия на соответствующие рецепторы), образующих каналы для Са2+.
Все эти механизмы могут приводить к усилению инфлюкса кальция из внеклеточной жидкости и его высвобождению из депо в цитоплазму клетки. Примеры веществ, повреждающих механизмы поддержания гомеостаза кальция внутри клеток, представлены в таблице 2.
Таблица 2. Некоторые вещества, нарушающие распределение кальция внутри клетки
| 1. Высвобождение кальция из митохондрий:- динитрофенол - динитрокрезол - хиноны - перекиси - аллоксан - железо - кадмий |
3. Усиление поступление кальция через плазматическую мембрану:- четыреххлористый углерод - диметилнитрозамин - парацетамол - 2,3,7,8-тетрахлордибензо-п-диоксин |
| 2. Высвобождение кальция из эндоплазматического ретикулума: - четыреххлористый углерод - бромбензол - хиноны - перекиси - альдегиды |
4. Угнетение эффлюкса кальция из клетки:- цистамин - хиноны - перекиси - дикват - ванадий |
Цитотоксический эффект в результате стойкого повышения содержания кальция в цитоплазме в настоящее время связывают с развивающимся при этом повреждением целостности цитоскелета и неконтролируемой активацией катаболических энзимов (фосфолипаз, протеаз, эндонуклеаз).
2.1. Повреждение цитоскелета
Цитоплазма клетки помимо цитозоля и клеточных органелл, как правило, содержит еще и нитевидные белковые структуры, которые в массе формируют клеточный скелет. Это образование выполняет не только стабилизирующую и структурирующую, но и другие функции, среди которых обеспечение клеточного деления, внутриклеточный транспорт, секреция, обмен рецепторных белков, регуляция клеточной подвижности и формы. Белки цитоскелета эпителиальных клеток формируют филаменты кератина, в мышечных клетках - десмин или волокна актина и миозина, в соединительнотканных клетках - виментин, в эритроцитах - спектрин, в нервных и глиальных клетках - нейро- и глиофиламенты. Важную роль в физиологии клеток играют микротрубочки, образованные молекулами тубулина, организованные в характерные нитевидные структуры. Они принимают участие в формировании митотического веретена делящихся клеток.
Велика роль структур, формирующих цитоскелет в нервных клетках. Здесь микрофиламенты и микротрубочки принимают участие в монтаже системы антероградного и ретроградного аксонального транспорта биологически активных веществ. Как известно белки синтезируются в перикарионе нейрона, а затем транспортируются с помощью этой системы на периферию клетки (в область синаптических контактов). Транспорт по аксону необходим и для обеспечения функций аксональных мембран: оборот глобулярных протеинов в мембране аксонов, выполняющих функции ионных каналов (см. выше). Состояние Швановских клеток также контролируется трофическими веществами, переносимыми с аксональным током. С аксональным током перемещается весь набор энзимов, нейромедиаторов и их предшественников, необходимых для обеспечения синаптической передачи нервных импульсов.
При действии разнообразных веществ на изолированные клетки (культура ткани) выявляется отчетливое изменение формы их поверхности: появляются выпячивания цитоплазмы, называемые пузырьками (blebs). Такое "пузырение" (или вскипание - blebbing) клеточной мембраны - один из ранних и надежных признаков разрушения сети цитоскелета. Вещества, нарушающие гомеостаз внутриклеточного кальция, инициируют "вскипание" мембраны. Механизм феномена объясняют следующим образом. Кальций вовлечен в процесс поддержания структуры цитоскелета как непосредственно, так и через ряд Са2+-связывающих протеинов и Са2+-зависимых энзимов. Особенно значим уровень Са2+ в цитоплазме для образования ассоциации белков цитоскелета с белками плазматической мембраны и взаимодействия различных элементов цитоскелета. Стойкое увеличение концентрации кальция в цитозоле по-видимому вызывает отщепление актина микрофиламентов от
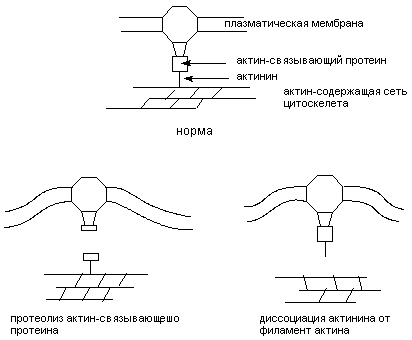
Рисунок 6. Са2+-зависимые механизмы повреждения цитоскелета клеток, обработанных хинонами
Белки цитоскелета повреждаются не только в результате повышения содержания Са2+ в цитоплазме, но и в результате воздействия токсикантов с иными механизмами токсического действия. Так, непосредственно взаимодействуют с белками цитохалазины, фаллоидин (один из токсинов бледной поганки), а также колхицин (рисунок 7), алкалоиды барвинка (винбластин, винкристин).
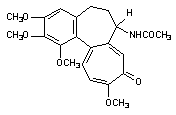
Рисунок 7. Структура колхицина
Действие веществ на тубулин митотического веретена делящихся клеток приводит к нарушению пролиферации последних (особенно клеток системы крови). Колхицин и алкалоиды барвинка в эксперименте, разрушая цитоскелет аксонов нервных клеток, существенно нарушают аксональный ток. Вероятно, аналогичные эффекты могут развиваться и при повреждении цитоскелета веществами, нарушающими внутриклеточный гомеостаз кальция.
2.2. Активация фосфолипаз
Фосфолипазы катализируют гидролиз фосфолипидов, входящих в состав клеточных мембран. Эти энзимы широко представлены в различных клетках. Особое внимание исследователи уделяют фосфолипазам А2 - группе липаз, основная функция которых состоит в удалении из мембраны поврежденных фосфолипидов, путем высвобождения жирных кислот, подвергшихся пероксидации. Фосфолипазы А2 являются Ca2+- и кальмодулин-зависимыми энзимами, и, следовательно, чувствительными к повышению кальция в цитоплазме. Стимуляция фосфолипазы кальцием приводит к усилению разрушения фосфолипидов мембран и повреждению клеток. Повышение содержания кальция в цитоплазме, активацию фосфолипазы, ускорение оборота фосфолипидов с последующей гибелью гепатоцитов регистрировали при токсическом поражении печени четыреххлористым углеродом. Аналогичные изменения наблюдали в миокардиоцитах и гепатоцитах, инкубируемых в условиях аноксии. Хлорпромазин, угнетающий активность фосфолипаз, защищает клетки от повреждающего действия циототоксикантов, нарушающих внутриклеточный гомеостаз кальция. Эти и другие данные подтверждают значение рассматриваемого механизма в развитии токсического повреждения клеток.
2.3. Активация протеаз
К числу протеаз, с оптимумом активности в области нейтральных значений рН, относятся: АТФ-зависимые, убиквитин-зависимые, Са2+_зависимые (кальпаины) протеазы. Кальпаины присутствуют практически во всех клетках млекопитающих. Они локализуются вне лизосом, в мембранных структурах в форме неактивного комплекса с ингибиторными протеинами (кальпастатины). Основные функции кальпаинов - репарация цитоскелета и клеточных мембран, разрушение рецепторных протеинов и их обновление, активация некоторых энзимов, участие в процессах митоза.
Идентифицированы две изоформы энзимов - с высоким и низким сродством к кальцию, обе активируемые этим ионом. В эксперименте показано, что стойкая, неуправляемая активация кальпаинов кальцием приводит к повреждению микрофиламентов цитоскелета тромбоцитов, клеточных мембран эритроцитов, гибели клеток печени, миокардиоцитов и т.д.
2.4. Активация эндонуклеаз
При завершении клеткой жизненного цикла активируется процесс "программированной" физиологической клеточной гибели - апоптоз. На ранних этапах в апаптотической клетке проявляются морфологические изменения: "вскипание" клеточной и ядерной мембран, конденсация хроматина. Самым надежным маркером развивающегося процесса является активация эндонуклеаз, энзимов, расщепляющих хроматин на фрагменты - олигонуклеосомы. Активация эндонуклеаз отмечается при гибели тимоцитов и лимфоцитов в облученном организме.
Установлено, что кальций участвует в активации эндонуклеаз. Вызванная глюкокортикоидами фрагментация ДНК тимоцитов и гибель этих клеток связаны со стойким повышением содержания Са2+ в цитоплазме. Одним из постоянных проявлений отравления экспериментальных животных диоксином является атрофия тимуса. Установлено, что при этом в тимоцитах значительно активируется процесс фрагментации ДНК, причем периоду разрушения хроматина предшествует период повышения концентрации кальция в цитоплазме. Возможно, что механизм цитотоксичности ТХДД хотя бы отчасти связан с кальциевой активацией эндонуклеаз.
Активация эндонуклеаз может быть причиной гибели клеток печени, миокарда, почек при отравлениях многими химическими веществами.
3. Активация свободно-радикальных процессов в клетке
3.1. Сущность явления
Многие ксенобиотики, попав во внутренние среды организма, подвергаются метаболическим превращениям (см. раздел "Биотрансформация ксенобиотиков в организме"). Одним из возможных результатов метаболизма является образование реактивных промежуточных продуктов. Взаимодействие этих реактивных метаболитов, либо вторичных продуктов их превращения, с молекулами-мишенями приводит к нарушению клеточных функций. Изменения в клетках могут быть следствием как избирательного повреждения какой либо одной биохимической структуры, так и сочетанного повреждения нескольких структурно-функциональных элементов. Достаточно часто в ходе исследований удается идентифицировать реактивный метаболит, изучить особенности его взаимодействия с молекулами-мишенями, оценить факторы облегчающие и модулирующие эти взаимодействия. Становиться все более очевидным, что многие реактивные промежуточные продукты метаболизма ксенобиотиков появляются в форме свободных радикалов, т.е. на внешней орбитали молекулы метаболита находится неспаренный электрон. Центром образования такого радикала в молекуле могут быть атомы углерода, азота, кислорода, серы (рисунок 8):
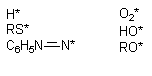
Рисунок 8. Примеры простых свободных радикалов
Хорошо доказана возможность образования радикалов при метаболизме ацетаминофена, адриамицина (и других цитостатиков антрациклиновой группы), нитрофурантиона, параквата, фенилгидразина, четыреххлористого углерода, бенз(а)пирена и т.д. (рис. 9).
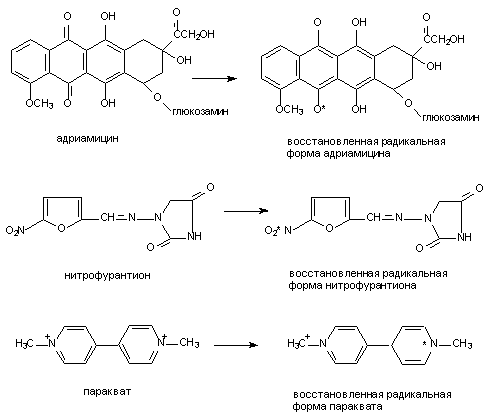
Рисунок 9. Структура веществ, подвергающихся биотрансформации с образованием активных радикалов
Появление неспаренного электрона на внешней орбитали делает метаболит чрезвычайно реакционноспособным. Активные радикалы вступают внутри клетки в многочисленные реакции, в том числе, связываются с ненасыщенными жирными кислотами, отщепляют водород от других молекул, взаимодействуют друг с другом с образованием димеров и т.д.
Свободные радикалы, такие как анионы семихинонов, азо-анионы, анионы нитроароматических соединенй, биспиридиниевые катионы, могут активировать молекулярный кислород путем одновалентного восстановления последнего до супероксид-аниона (О2-*). Супероксид при взаимодействии с водой с большой скоростью дисмутирует с образованием перекиси водорода (Н2О2) и чрезвычайно активного оксиданта - гидроксильного радикала (*ОН). Некоторые металлы с переменной валентностью (медь, железо) способны катализировать в организме реакции такого типа. Эти, так называемые, вторичные радикалы представляют высокую опасность для клетки. Обладая достаточной стабильностью, они взаимодействуют с самыми разными биомолекулами, и не только повреждают их, но и провоцируют цепные реакции дальнейшего образования активных радикалов из липидов, аминокислот, нуклеиновых кислот и т.д. Интегральный эффект такого каскада радикал-инициирующих реакций приводит к значительному нарушению физиологии клетки, её повреждению. На макроскопическом уровне это проявляется некрозом ткани, развитием фиброза в пораженных органах, а в отдаленный период - появлением новообразований (рисунок 10).

Рисунок 10. Активация свободно-радикальных процессов в клетках и их последствия
3.2. Механизмы клеточной антирадикальной защиты
Свободные активные радикалы в норме в клетке образуются постоянно. Так, в процессе метаболизма веществ в гладком эндоплазматическом ретикулуме флавопротеины, а в митохондриях окислительные энзимы цепи дыхательных ферментов, постоянно продуцируют некоторое количество супероксид-иона (О2-*) и перекиси водорода (Н2О2). Однако содержание в клетке этих и других радикалов жестко контролируются широким спектром биохимических инструментов антирадикальной защиты, включая супероксиддисмутазу, каталазу, GSH-пероксидазу, GSSG-редуктазу,
Механизмы антирадикальной защиты включают как ферментативные, так и неферментативные процессы. Самым простым примером некаталитического разрушения радикалов является их гидролиз, лежащий в основе нейтрализации многих водорастворимых продуктов, например, ацилгалидов, эпоксидов, карбокатионов, изоцианатов, эписульфониум-иона и т.д.
Наиболее важной неферментативной реакцией "обезвреживания" радикалов является их взаимодействие с биологическими антиоксидантами, такими как витамин Е, глутатион, витамин С. В результате такого взаимодействия образуются нереакционноспособные вещества, прерывание каскад "наработки" свободных радикалов.
Гомеостаз в клетке поддерживается за счет равенства скоростей образования и связывания радикалов. В случае повреждения механизмов защиты клеток, либо активации процессов образования радикалов, превосходящих по интенсивности возможности защиты, или даже разрушающих эти механизмы, развивается поражение клетки. Так, интоксикация преимущественным пульмонотоксикантом паракватом приводит к некоторому снижению содержания глутатиона в печени. Предварительное связывание глутатиона диэтилмалеатом приводит к тому, что паракват приобретает свойства преимущественного гепатотоксиканта. Таким образом, резерв глутатиона в клетке имеет особое значение для обеспечения её антиоксидантной защиты.
Хотя глутатион может взаимодействовать с многочисленными субстратами и неферментативно, наличие в тканях энзима глутатион-S-трансферазы (GST) значительно ускоряет течение процесса, повышает его эффективность. Множественность форм GST, их широкая субстратная специфичность, высокий уровень активности в различных тканях делают систему глутатионтрансфераз наиболее универсальной и значимой для связывания активных метаболитов.
Глутатион и селен-зависимые глутатионпероксидазы восстанавливают перекись водорода и другие гидроперекиси до менее токсичных алкоголей и воды. Глутатион-дисульфид, образующийся в ходе этой реакции, подвергается обратному восстановлению до глутатиона с помощью НАДФН-зависимой глутатионредуктазы. Активность глутатионредуктазы ингибируют изоцианат-содержащие продукты метаболизма нитрозомочевины.
Два других энзима, имеющих большое значение для детоксикации свободных радикалов, это супероксиддисмутаза (СОД) и каталаза. Первый из энзимов катализирует преобразование двух супероксидных радикалов в молекулу кислорода и перекись водорода. Обнаруживаемая во всех тканях СОД содержит в структуре активного центра ионы Cu, Zn, Mn. Образующаяся перекись водорода разрушается с помощью каталазы или глутатионпероксидазного цикла.
3.3. Механизмы активации ксенобиотиков и образования свободных радикалов
Ксенобиотики могут трансформироваться в радикалы как энзиматическим, так и неэнзиматическим путем. Например известны токсиканты, специфично повреждающие тела дофаминэргических и серотонинэргических нейронов ЦНС (6-гидроксидофамин и 5,7-дигидрокситриптпмин),
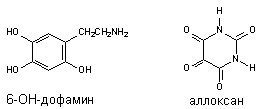
Рисунок 11. Вещества, инициирующие свободно-радикальные процессы путем спонтанного окислительно-восстановительного превращения молекулы в клетках
Ионы металлов с переменной валентностью (медь, железо) облегчают процесс аутоокисления, а восстановители, например аскорбиновая кислота, обеспечивают регенерацию исходной формы ксенобиотика. Таким образом, формируется неэнзиматический окислительно-восстановительный цикл токсиканта. SH-соединения, никотиновая кислота в опытах in vitro останавливают процесс.
Ультрафиолетовые лучи активируют превращение (фотоактивация) накапливающихся в коже сульфаниламидных препаратов, 4-аминобензойной кислоты (рисунок 12) и др. в свободно-радикальную форму, которая, как полагают, ответственна за развитие фототоксических и фотоаллергических процессов у лиц, принимающих эти лекарства.
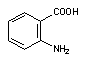
Рисунок 12. Структура о-аминобензойной (антраниловой) кислоты
При лечении псориаза псораленом и его аналогами одновременно назначаемое длинноволновое ультрафиолетовое облучение (320 - 400 нм) кожи пациентов активирует превращение препаратов в свободно-радикальную форму. Активные радикалы повреждают измененные эпидермальные клетки, образуя фотоаддукты с пиримидиновыми основаниями их ДНК. Наступает ремиссия болезни, но побочным неблагоприятным эффектом является развитие в последующем меланом и сквамозноклеточной карциномы, так как в процессе лечения изменяется генетический код и некоторой части здоровых эпидермоцитов.
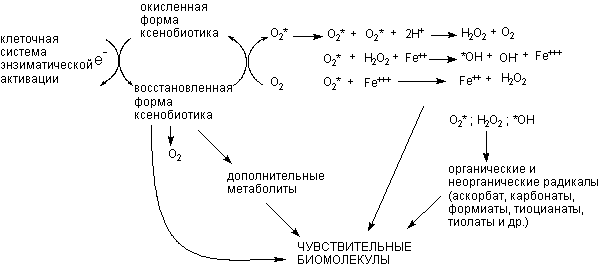 + Щелкните для загрузки увеличенной копии (5,08кб, 604x270 GIF) |
| Рисунок 13. Окислительно-восстановительный цикл трансформации ксенобиотиков, сопровождающийся активацией свободно-радикальных процессов в клетке. О2* - супероксидный анион; *ОН - гидроксильный радикал |
Однако основной путь образования свободных радикалов в клетке - энзиматический метаболизм ксенобиотиков (рисунок 13). Способность веществ метаболизировать с образованием радикалов обычно связывают с величиной их одноэлектронного восстановительного потенциала. Соединения с высоким сродством к электронам предрасположены к их акцепции и легко восстанавливаются биологическими системами, в то время как вещества с низким сродством к электрону восстанавливаются биосистемами плохо. Вещества, не вступающие в окислительно-восстановительный цикл не являются источниками образования свободных радикалов в клетках. Например, хлороформ (НССl3) является слабым источником прооксидантных процессов из-за низкой способности к оденоэлектронному восстановлению. Напротив, четыреххлористый углерод (CCl4) легко метаболизирует в трихлорметильный радикал (*ССl3), способный отнимать водородные атомы от ненасыщенных жирных кислот, и является инициатором перекисного окисления липидов. Кроме того *CCl3 связывается с липидами микросомальных мембран, активирует кислород, который в свою очередь взаимодействует с макромолекулами (белками, нуклеиновыми кислотами). Восстановление четырёххлористого углерода до трихлорметилового радикала катализируется комплексом оксидаз смешанной функции, состоящим из флавопротеинов, НАДФН-зависимой Р-450 релуктазы, НАДН-цитохром b5 редуктазы и оксидазы цитохрома Р-450.
В процессе метаболизма адриамицина, митомицина С, нитрофурантиона, параквата и некоторых других ксенобиотиков образование промежуточных свободных радикалов идет при участии одной лишь НАДФН-зависимой цитохром Р-450-редуктазы. При этом образовавшиеся радикалы удается выявить только в жестких анаэробных условиях. В присутствии кислорода восстановленные радикалы быстро окисляются до исходной формы, при этом электроны переходят с радикала на молекулу кислорода, что приводит к образованию супероксидного аниона и других его реактивных форм и также активации свободно-радикальных процессов.
Очевидно, что если такой химический окислительно-восстановительный цикл превращения ксенобиотика будет продолжаться в течение достаточно длительного времени, механизмы клеточной защиты могут истощиться и произойдет повреждение клетки. Такая возможность вполне вероятна, поскольку известно, что супероксид-анион инактивирует супероксиддисмутазу, превращает аскорбиновую кислоту и токоферол, а гидроксильный радикал - угнетает глутатионпероксидазу.
Помимо гладкого эндоплазматического ретикулума оксидазы смешанной функции обнаружены в мембране ядра клетки. Поскольку эта мембрана окружает хроматин, активация здесь ксенобиотиков и образование свободных радикалов представляет угрозу ДНК, либо вследствие прямого взаимодействия метаболитов с нуклеиновыми кислотами, либо опосредованно, путем образования реактивных метаболитов кислорода или продуктов перекисного окисления липидов - компонентов ядерной мембраны.
Помимо упомянутых выше, в образовании активных радикалов могут принимать участие и другие энзимы. Так, ксантиноксидаза участвует в метаболизме адриамицина, нитрофурантиола, параквата до продуктов их одноэлектронного восстановления. Тирозиназа, в большом количестве содержащаяся в клетках меланом участвует в образовании многих реактивных метаболитов. Цитоплазматическая диафораза и простогландинсинтетаза облегчают формирование окислительно-восстановительного цикла трансформации параквата, ацетаминофена, бенз(а)пирена и т.д.
Таким образом, можно выделить несколько ключевых моментов, имеющих особое значение для реализации повреждающего действия ксенобиотиков на клетку, путем активации свободно-радикальных процессов:
1. Образовавшись, радикал - промежуточный продукт может иметь несколько способов дальнейшего превращения, соотношение между которыми зависит от степени оксигенации клеток (тканей);
2. Усиленное образование свободных радикалов может начаться в нескольких независимых локусах клетки (эндоплазматическом ретикулуме, митохондриях, ядре, цитоплазме);
3. Активация ксенобиотиков до активных радикалов может стать следствием последовательного действия на токсикант нескольких ферментов;
4. Возможно неэнзиматическое образование свободных радикалов. Превращение одного из ксенобиотиков может активировать превращение другого. Так, блеомицин повреждает ДНК в присутствии митомицина С и т.д.
3.4. Биологические последствия активации свободно-радикального процесса в клетке
В результате образования свободных агрессивных радикалов повреждаются самые разные структуры-мишени: липидные мембраны, свободные аминокислоты, полисахариды, нуклеиновые кислоты, рецепторные молекулярные комплексы, транспортные протеины. Итогом такого действия является изменение функционального состояния и гибель клетки, мутация её генетического кода, что, как уже указывалось, на уровне макроорганизма приводит к явлению массивной клеточной гибели (некроз), разрастанию соединительной ткани в органе (фиброз), мутагенезу, развитию новообразований в отдаленные периоды после действия токсиканта.
Характер повреждающего действия активных радикалов во многом определяется их стабильностью и расстоянием, на которое они могут мигрировать от места своего образования. В соответствии с этими признаками реактивные метаболиты могут быть разделены на несколько групп.
Наивысшей реакционной способностью обладают промежуточные продукты, образующиеся в ходе так называемого "суицидного метаболизма". Эти продукты разрушают образующие их энзимы. К числу "суицидных" ингибиторов Р-450 относятся, в частности, винилгалогены, некоторые олифены, дигидропиридины, циклопропильные соединения.
Следующими по реакционной способности являются вещества, покидающие область локализации фермент-субстратного комплекса, но активно взаимодействующие с другими протеинами. Примерами веществ, действующих подобным образом, являются: ацилгалидный метаболит хлорамфеникола, электрофильный радикал серы, отщепляющийся в ходе метаболизма паратиона и т.д.
Менее реакционноспособными являются продукты, мигрирующие в другие локусы клеток и даже за их пределы. Эта группа метаболитов включает большинство промежуточных продуктов. Подобные радикалы способны проникать через биологические мембраны. Это свойство присуще метаболитам бенз(а)пирена, бромбензола, диметилнитрозамина, винилхлорида, трихлорэтилена и т.д.
Еще меньшей раекционной способностью обладают метаболиты, способные выходить за пределы органов, в которых они образуются, и повреждають другие органы и ткани. Примерами этой группы метаболитолв являются: метаболит гексана - 2,5-гександион, вызывающий периферическую нейропатию, пирролизидиновых алкалоидов - пирролы, мигрирующие из печени в легкие, где действуют на эндотелий сосудов, эпоксиды 4-винилциклогексана, вызывающие поражение яичников.
Механизмы токсического действия свободных радикалов представлены на рисунке 14.
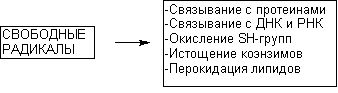
Рисунок 14. Некоторые токсические эффекты свободных радикалов
4. Повреждение мембранных структур
4.1. Основные свойства и функции биологических мембран
1. Плазматическая мембрана. Содержимое клетки отграничено от окружающей среды плазматической мембраной. Благодаря этому поддерживаются условия, позволяющие структурным элементам клетки выполнять присущие им функции. С другой стороны через плазматическую мембрану осуществляется непрерывный транспорт различных молекул, чем и обеспечивается обмен веществом между средой и клеткой. Например плазматическая мембрана печеночной клетки имеет площадь около 8000 мкм2. Такая большая площадь контакта с окружающей средой полностью обеспечивает обмен веществ клетки. Толщина плазматической мембраны составляет около 10 нм. Наконец в мембране локализуются белковые структуры, формирующие ионные каналы, рецепторы для физиологически активных веществ, обеспечивающие восприятие сигналов, регулирующих биологическую активность клетки (см. ниже).
Мембрана образована двумя слоями молекул липидов, гидрофобные части которых направлены друг к другу, а гидрофильные в сторону окружающей и внутренней среды клетки. Основные группы липидов - фосфолипиды (фосфатидилхолин, фосфатидилэтаноламин, фосфатидилсерин, сфингомиелин), гликолипиды, нейтральные липиды (холестерол и т.д.). Молекулы липидов легко диффундируют в липидных слоях. Время нахождения отдельной молекулы на неизменной позиции составляет 10-8 - 10-9 сек. Обмен молекулами между слоями осуществляется редко. Распределение молекул различного строения между слоями асимметрично.
В мембрану встроены белковые молекулы, которые часто пронизывают всю ее толщу, либо погружаются на различную глубину, локализуясь на внешней или внутренней стороне. Углеводный компонент клеточной мембраны представлен главным образом гликопротеинами. Они располагаются на внешней поверхности мембраны. Мембраны различных клеток существенно различаются по своему строению и функциям. В клеточной мембране печени обнаруживается около 20 энзимов. Активность энзимов во многом зависит от липидного состава мембран. Особыми свойствами обладают возбудимые мембраны нервной, мышечной и железистой тканей. Изменение их свойств в ответ на стимул являются важнейшим звеном цепи биологических процессов, лежащих в основе формирования реакции организма на внешние и внутренние раздражители.
Плазматические мембраны, несмотря на преимущественно липидный состав, способны пропускать низкомолекулярные водорастворимые вещества, так как содержат поры со средним диаметром 0,7 - 1,0 нм. Полагают, что поры, заполненные водой, сформированы белковыми молекулами. Объем воды, содержащейся в мембранах, составляет от 30 до 50% от общего их объёма. Максимальная молекулярная масса водорастворимого вещества, способного проникать через плазматические мембраны, - около 200 дальтон.
Клеточные мембраны чрезвычайно динамичный элемент. Их строение изменяется в соответствии с условиями окружающей среды и потребностями клетки. Увеличение площади поверхности растущей клетки осуществляется за счет слияния клеточной мембраны с синтезируемыми в цитоплазме (аппарат Гольджи) микровезикулами. Этот же процесс обнаруживается и у не растущих секреторных и нервных клеток, которые путем отшнуровывания микровезикул выделяют в окружающую среду биологически активные вещества (гормоны, нейромедиаторы).
Известен и противоположный процесс - поступления необходимых веществ из окружающей среды в клетку, путем захвата фрагментом клеточной мембраны субстрата, погружением его в цитоплазму и отшнуровывания от остальной мембраны. Слияние мембраны с везикулами и их отшнуровывание обеспечивают динамичность её состава.
Важными свойствами клеточной мембраны является её электрический заряд и электрическая проводимость. Наружная сторона мембраны клеток в состоянии покоя заряжена положительно. Полярность мембраны определяется отчасти асимметричностью липидных слоев, отчасти наличием в её составе белковых молекул и гликопротеинов. Особое значение имеет градиент концентрации ионов по обе стороны мембраны, поддерживаемый благодаря энергозатратным процессам. Таким образом свойства мембраны и обмен веществ в клетке тесно связаны. Вещества, вмешивающиеся в обмен липидов, существенно влияют на свойства биологических мембран.
2. Цитоплазматические мембраны. Внутри клеток имеются многочисленные мембранные структуры, образующие эндоплазматический ретикулум, мембрану ядра клетки, митохондрии, аппарат Гольджи, лизосомы. Эти структуры образованы двумя слоями биологической мембраны. Между этими слоями имеется пространство толщиной около 10 нм. Общая толщина внутриклеточных мембран - около 25 нм.
Мембрана клеточного ядра представляет собой специализированный отдел эндоплазматического ретикулума. Пространство, находящееся между двумя листками мембраны, по-видимому сообщается с межклеточным пространством. Этим путем токсиканты могут проникать непосредственно в ядро, минуя цитоплазму, что прослеживается, в частности, при поступлении в клетку акридиновых красителей. Ионы Na+ также поступают в ядро из экстрацеллюлярной жидкости по эндоплазматическому ретикулуму. Наличие пор в ядерной мембране диаметром 50 - 100 нм обеспечивает обмен веществом между ядром и цитоплазмой. Даже макромолекулы (РНК, энзимы и др.) проходят через мембрану путем простой диффузии.
Цитоплазматические мембраны служат организующим субстратом для объединение в единый комплекс ряда энзиматических систем. Например, мембраны митохондрий представляют собой сложную мозаику взаимодействующих энзиматических групп. Складчатость мембран способствуют увеличению площади их поверхности и тем самым интенсификации обменных процессов. Общая площадь мембран митохондрий печеночной клетки оценивается в среднем в 29000 мкм2. В качестве матрикса для энзимов выступают и другие мембранные структуры: гладкий и шероховатый эндоплазматический ретикулум, аппарат Гольджи. Эти структуры участвуют в обмене веществ. Так, на рибосомах, локализующихся на мембранах шероховатого ретикулума, осуществляется синтез белка; энзимы гладкого эндоплазматического ретикулума - важнейший элемент метаболизма ксенобиотиков; аппарат Гольлджи - основная секреторная структура клетки.
Токсическое действие многих веществ сопряжено с их влиянием на состояние мембранных структур. Оно может быть прямым и опосредованным. Наиболее вероятными механизмами опосредованного повреждения биологических мембран при интоксикациях являются:
- активация перекисного окисления липидов;
- активация фосфолипазной активности.
4.2. Действие токсикантов на мембраны
4.2.1. Прямое действие на мембраны
Возможно действие ксенобиотиков непосредственно на липидный бислой или белковые компоненты биомембран. Развивающиеся вследствие этого эффекты могут быть как неспецифичными (отсутствие строгой зависимости между структурой токсиканта и развивающимся эффектом), так и строго специфичными. Специфичность является следствием действия токсикантов на энзимы и рецепторные белковые комплексы (см. выше). Часто действие носит смешанный "полуспецифический" характер.
В результате неспецифического действия многочисленных токсикантов (бензола, толуола, динитробензола, хлороформа, мылов, сапонинов, смачивающих веществ, тяжелых металлов и других денатурирующих агентов) может нарушаться структурная целостность мембран, что приводит к деформации, лизису клетки и её гибели. При действии таких веществ на мембраны эритроцитов развивается гемолиз.
Действуя в малых дозах на возбудимые мембраны нервных клеток ЦНС, органические растворители, спирты вызывают седативно-гипнотический. В действии этих соединений, помимо неспецифических, прослеживаются и отчетливые специфические механизмы. Так, вещества со свойствами "неэлектролитов" накапливаясь в мембранах, одновременно изменяют их проницаемость для ионов Na+, K+, Ca+2, Cl- (рис. 15).
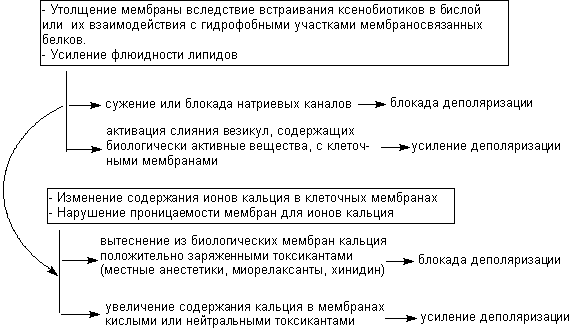 + Щелкните для загрузки увеличенной копии (7,4кб, 571x331 GIF) |
| Рисунок 15. Возможные механизмы непосредственного действия токсикантов на биологические мембраны |
Многие токсиканты избирательно действуют на мембраносвязанные энзимы, транспортные системы, рецепторные комплексы к биологически активным веществам. К числу таких в частности относятся гликозиды, агонисты и антагонисты нейромедиаторов и гормонов, блокаторы ионных каналов и многие другие.
4.2.2. Активация перекисного окисления липидов
Благодаря ненасыщенности углеводородной цепи жирных кислот и непрочности связи аллильных атомов водорода с метиленовыми группами, фосфолипиды клеточных мембран наиболее предрасположены к реакции окисления, инициируемой свободными радикалами, образующимися в клетке. Этому способствует и то обстоятельство, что молекулярный кислород в 7 - 8 раз лучше растворяется в липидной фазе, чем в воде и гидрофильных сайтах клетки. Существенная активация процесса образования свободных радикалов при химических воздействиях приводит к усилению перекисного окисления липидов и повреждению биологических мембран. Процесс перекисного окисления усиливается вторичным образованием из липидов высоко реакционно-способных и легко диффундирующих карбонильных радикалов, которые могут обусловливать неблагоприятные процессы, развивающиеся в клетках далеко за пределами сайта своего образования не только клетки, но и органа.
Не смотря на многочисленные исследования до настоящего времени нет четких представлений о значении активации перекисного окисления липидов для развития токсического процесса, инициируемого даже такими мощными прооксидантами, как паракват, адриамицин, нитрофурантион и т.д.
Для развития патологического процесса важно, чтобы окислительно-восстановительный цикл превращения ксенобиотика "работал" достаточно долго, с тем, чтобы произошло истощение механизмов антирадикальной защиты клетки (см. выше). Поскольку непродолжительная инициация окислительно-восстановительного цикла трансформации ксенобиотика может стимулировать систему антиокисдантной защиты, при интоксикациях некоторыми прооксидантами возможно развитие парадоксальной реакции, а именно - угнетение процесса окисления липидов в тканях.
Для развития процесса важна степень насыщенности тканей кислородом. В полностью аэробной среде образующийся из ксенобиотика восстановленный метаболит подвергается быстрому обратному окислению. При этом значительно возрастает количество радикалов в виде вторичных активных форм кислорода. Поэтому хорошо снабжаемые кровью и насыщенные кислородом ткани (легкие, сердце, головной мозг) являются более чувствительными к повреждающему действию прооксидантов.
4.2.3. Активация фосфолипаз
Важным механизмом повреждения биологических мембран является гидролиз фосфолипидов, наступающий вследствие активации фосфолипаз (особенно фосфолипазы A2). Активация энзима происходит в результате прямого или опосредованного (гипоксия, нарушение гомеостаза внутриклеточного кальция и т.д.) действия многих токсикантов на клетки организма (рисунок 16).
Результатом действия фосфолипазы А2 на липиды биологических мембран является высвобождение арахидоновой кислоты. Последняя является, в свою очередь, субстратом энзима циклооксигеназы. Превращение арахидоновой кислоты под влиянием энзима приводит к образованию эйкозаноидов (простагландинов, тромбоксанов, простациклинов) - веществ, активирующих воспалительные процессы в тканях. Под влиянием другого энзима, 5-липоксигеназы, арахидоновая кислота превращается в лейкотриены и эйкозатетраеноевые кислоты. Эти вещества являются химиоатрактантами нейтрофилов, веществами, регулирующими сосудистую проницаемость.
Еще одним продуктом энзиматического расщепления липидов мембран является фактор агрегации тромбоцитов (ФАТ) - клон биологически активных веществ (более 150 аналогов) близкого строения. ФАТ чрезвычайно токсичные вещества (LD50 для кролика - 0,005 мг/кг; для собаки - 0,07 мг/кг), вызывающие при внутривенном введении шокоподобное состояние (острый, некупируемый коллапс, бронхоспазм и др.).
 + Щелкните для загрузки увеличенной копии (8,23кб, 595x576 GIF) |
| Рисунок 16. Образование биологически активных веществ при участии фосфолипаз, активируемых в ходе токсического процесса |
4.3. Биологические последствия действия токсикантов на мембраны
Вещества, действующие на клеточные мембраны, как правило, изменяют следующие их параметры:
1. Проницаемость. Изменяется поступление субстратов в клетки и отток продуктов метаболизма. Действие проявляется изменением интенсивности обмена веществ в клетке, нарушением электрических свойств мембран.
2. Электрический заряд. Нарушается раздражимость возбудимых клеток.
3. Структурная организация. Нарушается структура клетки, возможна клеточная гибель.
Кроме того, патохимические процессы, проходящие в мембране, приводят к образованию целого ряда биологически активных веществ (см. выше), участвующих в патогенезе токсического процесса.
Поскольку система внутриклеточных мембран также является мишенью для действия многочисленных токсикантов, среди них можно выделить группу митохондриальных ядов, повреждающих различные звенья процессов биоэнергетики, веществ действующих на шероховатый (нарушение процессов синтеза белка) и гладкий эндоплазматический ретикулум (индукция или угнетение метаболизма ксенобиотиков), лизосомальные мембраны (аутолиз клеток) и др.
5. Повреждение процессов синтеза белка и клеточного деления
Деление, рост, дифференциация клеток, их мутация и малигнизация - процессы, неразрывно связанные с обменом нуклеиновых кислот (ДНК и РНК) и синтезом белка. Эти процессы чрезвычайно сложны и включают множество этапов. Действие веществ, нарушающих нуклеиновый обмен и белковый синтез, в этой связи, весьма разнообразно (таблица 3). Подавляющее большинство механизмов токсического повреждения изучено в опытах in vitro на изолированных быстро размножающихся клетках, а порой даже на прокариотах. Вот почему среди приводимых в качестве примеров веществ, преобладают цитостатики, антибиотики и красители.
Таблица 3. Возможные точки приложения повреждающего действия токсикантов на процессы синтеза белка и нуклеиновых кислот.
1. Синтез ДНК. Репликация - изменение структуры (конформации) ДНК - нарушение процесса полимеризации ДНК - нарушение синтеза нуклеотидов - разрушение ДНК - нарушение процесса репарации ДНК - нарушение механизмов регуляции синтеза ДНК 2. Синтез РНК. Транскрипция - нарушение полимеризации РНК - нарушение процессии РНК - нарушение синтеза нуклеотидов - разрушение РНК - нарушение механизмов регуляции синтеза РНК 3. Синтез белка. Трансляция - нарушение организации и процессии рибосом и полисом - нарушение полимеризации аминокислот - нарушение образования аминоацетил-tРНК - нарушение формирования конформации белка и его третичной и четвертичной структур - нарушение механизмов регуляции трансляции |
5.1. Синтез ДНК. Репликация
Как указывалось ранее, нарушение репликации может явиться следствием химического взаимодействия ДНК с токсикантами. Однако возможны и иные механизмы повреждения (таблица 4).
Таблица 4. Механизмы действия ингибиторов синтеза нуклеиновых кислот и белков
>Механизм действия |
>Примеры |
| Ковалентное связывание с ДНК | иприт, циклофосфан |
| Индукция однонитевых разрывов | этиленоксид, этиленимин |
| Интеркалация | профлавин, этидиумбромид |
| Нековалентное связывание с ДНК | актиномицин |
| Взаимодействие с РНК-полимеразой | |
| Взаимодействие с ревертазой | стрептоварицин |
| Взаимодействие с ДНК-полимеразой | эдеин, налидиксовая кислота |
| Влияние на синтез нуклеотдиов | фаллоидин, цитозин-арабинозид |
| Нарушение обмена фолиевой кислоты, пиримидина, пурина | аминоптерин, аминопурин, азогуанин, азоцитозин |
| Нгнетение образования аминоацетил-tРНК | 6-флюоротриптофан, триптазан, порвалин |
1. Угнетение активности ДНК-полимераз и других ферментов синтеза.
Эдеин и налидиксовая кислота угнетают активность ДНК-полимераз, а N-диметилрифампицин подавляет РНК-зависимый синтез ДНК (ингибитор ревертазы). Соединения, нарушающие конформацию ДНК, могут подавлять реакцию её полимеризации, не изменяя активность соответствующих энзимов.
2. Влияние на синтез дезоксирибонуклеотидов.
Элементарной единицей ДНК являются дезоксирибонуклеотидтрифосфаты. Их синтез осуществляется в организме и может быть прерван на различных этапах (синтез пуринов, пиримидинов, нуклеотидов). Вещества, нарушающие синтез нуклеотидов, называются антиметаболитами. В качестве антиметаболитов могут выступать различные аналоги естественных предшественников нуклеотидов, которые вытесняют их из связи с энзимами. Частично эти аналоги перерабатываются и встраиваются в структуру нуклеиновой кислоты (2-аминопурин, 5-флюороурацил), при этом нарушается строение и функция последней. Некоторые вещества (нуклеозидные антибиотики - кордицепин) блокируют встраивание дезоксирибонуклеотидов в молекулу ДНК.
3. Влияние на регуляцию синтеза ДНК.
В качестве регуляторов репликации, у эукариотов выступают цитоплазматические факторы - специальные белки, ответственные за процесс инициации. Их проникновение в ядро показано экспериментально. Определенную роль играют также гуморальные факторы роста, гормоны. Так, кортикоиды и глюкагон усиливают синтез ДНК.
4. Действие на разрушение ДНК и её репарацию.
Эндонуклеазы и лигазы - энзимы, участвующие в репарации мутированных или неправильно синтезированных молекул ДНК. Так, поврежденный нуклеотид может быть вырезан из молекулы ДНК (эндонуклеазы), а на его место встроен неповрежденный фрагмент (лигазы). Резкое повышение внутриклеточной концентрации Са2+, наблюдающееся при острой интоксикации некоторыми токсикантами (см. выше), сопровождается активацией эндонуклеаз и разрушением ДНК.
Ингибиторы лигаз угнетают процессы репарации ДНК и повышают частоту мутаций (например, бензамид и его производные).
5. Другие механизмы действия.
Помимо повреждения ДНК и ферментов, участвующих в её синтезе и репарации, существуют и другие способы токсического нарушения процесса репликации. Так, в фазе приготовления к митозу возможно повреждение центриолей и синтеза митотического аппарата, образующего клеточное веретено. Веретено формируется SH-содержащими протеинами, которые благодаря -S-S- связям образуют нитевидные структуры. Естественно, токсиканты, взаимодействующие с SH-группами способны повреждать митотические веретена. Примерами таких токсикантов являются мышьяк, ртуть и их соединения, колхицин, подофилотоксин и др.
5.2. Синтез РНК. Транскрипция
В клетках определяются несколько форм РНК. Особое функциональное значение имеют: м-РНК, как матрица для синтеза полипептидов, t-РНК - молекула-переносчик аминокислот на рибосомы для их последующей полимеризации, r-РНК - составная часть рибосом. Возможны следующие механизмы повреждения РНК:
1. Нарушение конформации и свойств ДНК (см. выше).
На поврежденной ДНК синтезируется измененные молекулы РНК, не свойственные организму.
2. Угнетение активности РНК-полимераз.
Ингибитором РНК-полимеразы является, в частности, аманитин - яд бледной поганки.
3. Влияние на РНК-процессинг.
Это явление изучено недостаточно. Полагают, что кордицепин (см. выше) ингибирует начало процесса синтеза РНК.
4. Угнетение синтеза нуклеотидов.
Как уже указывалось, целый ряд веществ, так называемых антиметаболитов, способен угнетать синтез пуриновых и пиримидиновых оснований и нуклеотидов. Такие вещества, прежде всего, нарушают обмен РНК, находящихся в состоянии активного обращения. Возможно извращение синтеза РНК вследствие встраивания в молекулу "ложных" нуклеотидов (5-бромурацил, 8-азагуанин).
5. Нарушение регуляции синтеза РНК.
Синтез РНК контролируется целым рядом факторов. Большая часть генома находится в репримированном состоянии, т.е. блокирована специальными белками (гистонами) и не участвует в процессе транскрипции. Активация этих отрезков молекулы ДНК осуществляется низкомолекулярными хромосомными РНК, цитоплазматическими протеинами и другими факторами (например гормонами). Так, стероиды (эстрогены, андрогены, кортикостероиды) стимулируют синтез РНК и, следовательно, белка. Аналогичные эффекты описаны для инсулина и глюкагона. Многочисленные синтетические соединения (барбитураты, толбутамид, бензпирен, метилхолентрен, диоксин и т.д.) вызывают индукцию синтеза микросомальных энзимов, ответственных за метаболизм ксенобиотиков. По всей видимости, они взаимодействуют с репрессорными протеинами, вызывают депримирование ДНК и активируют тем самым синтез РНК и соответствующих белков.
5.3. Синтез белков. Трансляция
Объединение аминокислот в полипептидную цепь осуществляется на рибосомах или полисомах. Токсиканты могут вмешиваться в процессы синтеза белка и на этом этапе. Возможны следующие механизмы токсического действия:
1. Нарушение конформации рибосом и организации полисом.
Этот механизм экспериментально показан для целого ряда антибиотиков, действующих на рибосомы и полисомы бактериальных клеток.
2. Угнетение процесса образования полипептидной цепи.
Многие антибиотики вмешиваются в процесс синтеза белка на рибосомах бактерий. При этом выявляются нарушения:
- процессов считывания информации с кода мРНК;
- образования формилметионил-tРНК и повреждение тем самым инициации синтеза полипептидной цепи;
- образования пептидных связей (таков, в частности, один из механизмов токсического действия дифтирийного токсина);
- затруднение транслокации пептидил-tРНК и мРНК.
3. Угнетение синтеза и формирования аминоацил-tРНК.
Необходимым этапом полимеризации аминокислот и образования полипептидной цепи является их активация, путем образования комплекса с tРНК (образование аминоацил-tРНК). На первом этапе происходит взаимодействие аминокислот с АМФ (активация), и лишь затем образуется связь с РНК. Введение в среду, где инкубируется экспериментальный материал (делящиеся клетки, бактерии), ненормальных аминокислот, например D-форм или таких как этионин, тиенилаланин, алкилвалин и др., приводит к угнетению активности синтезирующих энзимов, а следовательно и процесса синтеза белка в целом.
4. Нарушение конформации синтезируемой белковой молекулы.
Синтезированная полипептидная цепь спонтанно формирует свою третичную структуру. Отчасти процесс проходит при участии различных кофакторов. Угнетение синтеза таких кофакторов или поступление в организм их неактивных аналогов (антивитаминов) нарушает процесс.
5. Влияние на регуляцию процесса трансляции.
Имеются данные о том, что процесс трансляции, особенно его инициация и терминация, подлежат контролю. Отвечают за это особые виды регуляторных РНК и протеинов, а также механизмы контролирующие уровень цАМФ в клетках. Образованию полисом способствует повышение содержания эстрадиола; инсулин активирует процесс инициации синтеза белка на малых субъединицах рибосом. Витамин К блокирует на уровне трансляции образование протромбина и других факторов свертывания крови и т.д. Усиление или угнетение рассмотренных процессов может лежать в основе токсического действия некоторых веществ.
5.4. Биологические последствия действия токсикантов на нуклеиновый обмен и синтез белка
Токсикологическое значение веществ, вмешивающихся в процесс синтеза белка и клеточное деление, обусловлено их цитостатическим, иммуносупрессорным, мутагенным, тератогенным и канцерогенным действием (см. соответствующие разделы).
Повреждающее действие химических веществ на ДНК называется генотоксическим. Наиболее чувствительны к генотоксическому действию клетки, способные к делению (эмбриональные, герменативные, костного мозга, эпителия почек, кожи, слизистой желудочно-кишечного тракта и т.д.). Последствия повреждения ДНК зависят от дозы токсиканта. Высокие дозы вызывают цитостатический эффект (гибель пула делящихся клеток), более низкие - канцерогенное, тератогенное, мутагенное действие. В основе канцерогенного, тератогенного, мутагенного действия лежат по сути общие механизмы, однако превращение конкретного вещества в канцероген, тератоген, мутаген зависит от целого ряда условий (таблица 5).
Таблица 5. Условия воздействия генотоксиканта, определяющие форму развития токсического процесса: канцерогенеза, мутагенеза, тератогенеза
Токсический процесс |
Чувствительная ткань |
Оптимальное время воздействия |
Продолжи тельность действия и доза |
Канцерогенез |
Любая пролиферирующая ткань | Неопределеное; любая стадия митоза | Обычно хроническое, беспороговое |
Мутагенез |
Герменативные клетки | Все стадии гаметогенеза | Острое и хроническое, беспороговое |
Тератогенез |
Все зародышевые ткани | Наивысшая - на ранних стадиях дифференциации тканей | Только острое, в дозах выше пороговых |
Существует представление, согласно которому проникновение в организм даже единственной молекулы генотоксиканта (в отличие от токсикантов с иным механизмом токсического действия) может привести к пагубным последствиям. Дело в том, что химическое повреждение единичной молекулы ДНК в единичной клетке макроорганизма, при стечении обстоятельств, может стать причиной мутогенеза, тератогенеза, канцерогенеза. Вероятность такого события бесконечно мала, но теоретически возможна. Такой характер действия веществ на биосистемы называется беспороговым.
Другим, важным с позиции токсикологии, последствием действия химических веществ на процессы синтеза белка является индукция микросомальных ферментов и, следовательно, изменение скорости и характера метаболизма ксенобиотиков. Значение этого эффекта неоднозначно, поскольку в итоге токсичность одних веществ уменьшается, других - возрастает (см. раздел "Метаболизм ксенобиотиков").
<< Содержание |