|
|
 |
|
ТОМ 3, СТ. 41 (стр. 278) // Ноябрь, 2002 г.
ТКАНЕСПЕЦИФИЧЕСКОЕ ДЕЙСТВИЕ ПЕПТИДОВ В КУЛЬТУРЕ ТКАНЕЙ КРЫС РАЗНОГО ВОЗРАСТА
"Успехи геронтологии", 2002г., выпуск 9
В.Х. Хавинсон, В.В. Малинин, Н.И. Чалисова, Е.И. Григорьев
Санкт-Петербургский институт биорегуляции и геронтологии СЗО РАМН,
197110, Санкт-Петербург, пр. Динамо, 3,
E-mail:vvm@medport.ru
Институт физиологии им. И.П. Павлова РАН,
199034, Санкт-Петербург, набережная Макарова, 6
Исследовано влияние полипептидных препаратов, выделенных из коры головного мозга (кортексин®), эпифиза (эпиталамин®), печени (гепалин®), тимуса (тималин®) и синтетических коротких пептидов - кортагена®, эпиталона®, ливагена®, вилона® на развитие эксплантатов коры и подкорковых структур головного мозга, печени, тимуса крыс разного возраста. Установлено, что исследуемые пептиды в зависимости от используемых концентраций оказывают тканеспецифическое действие в органотипических культурах тканей животных.
Изучение механизмов тканеспецифического действия пептидов является актуальной задачей современной молекулярной биологии, физиологии и медицины. Необходимо отметить, что понятие тканеспецифичности пептидов в научной литературе в настоящее время не имеет однозначного толкования. Вероятно, это связано с различными подходами к решению данной проблемы и особенностью интерпретации результатов исследований. Чаще всего под тканеспецифическим действием пептидов (также как и других веществ или воздействий) подразумевают механизмы их влияния на молекулярно-генетические или биохимические маркеры специализированных клеток-мишеней различных органов или тканей [5, 15, 16, 23, 24, 25].
Тканеспецифичность пептидов рассматривают также с точки зрения структурной организации белков различного происхождения. Установлено, что фракции низкомолекулярных компонентов экстрактов тканей характеризуются стабильным набором веществ пептидной природы, что позволяет считать определенные группы тканеспецифических пептидов характеристикой соответствующей ткани [17, 18]. В данном случае основной функцией тканеспецифических пептидов вероятнее всего является контроль пролиферации, дифференцировки и элиминации клеток соответствующей ткани. Примечательно, что действующее начало при этом представлено не отдельными компонентами, а множеством пептидов с разнонаправленной активностью.
В соответствии с предложенной ранее концепцией пептидной биорегуляции было выдвинуто предположение об участии эндогенных пептидов в поддержании структурного и функционального гомеостаза клеточных популяций, которые содержат и продуцируют эти факторы [4]. Принципиально важным в развитии представлений о механизмах тканеспецифического действия пептидов является исследование структурно-функциональных особенностей этих веществ. Известно, что тимоген (Glu-Trp) - синтетический аналог иммуноактивного дипептида, выделенного из тималина, оказывает стимулирующее действие на все этапы дифференцировки Т-лимфоцитов, индуцирует экспрессию дифференцировочных антигенов этих клеток, нормализует количество и соотношение Т- и В-лимфоцитов в крови при патологических состояниях. Вместе с тем, установлено, что активность тималина не ограничивается наличием пептидной фракции, послужившей основой для создания тимогена, а определяется также иммуноактивными фрагментами с другой аминокислотной последовательностью [6].
Необходимо подчеркнуть, что процесс выделения индивидуальных пептидов и определения их биологической активности является чрезвычайно трудоемким и длительным, требует оценки активности многих сотен пептидов, что практически трудно осуществимо в течение короткого периода времени. В связи с этим, в Санкт-Петербургском институте биорегуляции и геронтологии СЗО РАМН был разработан принципиально новый подход к поиску и синтезу физиологически активных пептидов [8]. В основе данного способа лежит аминокислотный анализ комплексных полипептидных препаратов, определение наиболее тканеспецифически значимых аминокислот для каждого препарата и далее конструирование первичной структуры пептида, учитывая данные расчетов энергетически выгодных конформаций для ионизированной молекулы. Такой подход позволил разработать технологию получения пептидных регуляторов функциональной активности тимуса, коры головного мозга, эпифиза, сетчатки, сосудов, сердца, бронхов, простаты и печени.
Результаты многолетних экспериментальных и клинических исследований полипептидных препаратов неидентифицированной структуры и синтетических коротких пептидов свидетельствуют как о полифункциональности, так и тканеспецифичности действия этих веществ [1, 3, 6, 7, 10, 12]. Однако, действие пептидов, особенно коротких, на развитие специализированных органов и тканей в настоящее время недостаточно изучено, что связано, в основном, с отсутствием адекватных экспериментальных моделей.
Материал и методы исследования
Наиболее удобным объектом для биотестирования пептидов являются органотипические культуры тканей, в которых отсутствуют нервные, гуморальные и другие воздействия на исследуемые клетки. Эксперименты, проведенные ранее [9, 11, 13], свидетельствуют о том, что природные и синтетические пептидные препараты оказывают тканеспецифическое действие на рост эксплантатов соответствующих тканей эмбрионов цыплят, 1- и 21-сут крыс. Несмотря на установленные факты, наиболее важным представляется изучение влияния пептидов на рост эксплантатов тканей животных различного возраста, в связи с существенными возрастными изменениями уровня синтеза белка и компенсаторно-приспособительных механизмов клеток.
В представленной работе исследовано влияние полипептидных препаратов, выделенных из коры головного мозга (кортексин), эпифиза (эпиталамин), печени (гепалин), тимуса (тималин) и синтетических коротких пептидов - кортагена, эпиталона, ливагена, вилона на развитие эксплантатов коры и подкорковых структур головного мозга, печени, тимуса крыс в возрасте 1 сут, 21 сут и 24 мес. Кортаген (Ala-Glu-Asp-Pro), эпиталон (Ala-Glu-Asp-Gly), ливаген (Lys-Glu-Asp-Ala) получены путем целенаправленного конструирования на основе анализа аминокислотного состава кортексина, эпиталамина и гепалина, соответственно [8]. Вилон (Lys-Glu) - синтетический пептид, созданный на основе фрагмента, встречающегося в составе эндогенных факторов тимуса и цитокинов [6, 21]. Эксперименты проведены на 900 эксплантатах различных тканей. Методика культивирования подробно описана ранее [11]. Исследуемые пептиды добавляли в культуральную среду в концентрациях от 2 до 200 нг/мл. Эффективными концентрациями для кортексина и гепалина были 100 нг/мл, для эпиталамина - 50 нг/мл, для тималина - 5 нг/мл, для синтетических пептидов - кортагена, эпиталона, ливагена - 20 нг/мл, за исключением вилона, у которого эффективная концентрация была 2 нг/мл. Контролем служили эксплантаты без добавления пептидов в культуральную среду. Чашки Петри с эксплантатами тканей помещали в термостат при температуре 37о С и через 3 сут просматривали в фазово-контрастном микроскопе. Затем определяли индекс площади (ИП), который рассчитывали в условных единицах как соотношение площади всего эксплантата, вместе с зоной выселяющихся клеток, к исходной площади эксплантата. Значения ИП выражали в процентах, контрольное значение ИП принимали за 100 %. Достоверность различий ИП в контрольных и опытных образцах оценивали с помощью t-критерия Стьюдента.
Результаты и обсуждение
Установлено, что в 1-е сут культивирования происходило распластывание экспланта-тов на коллагеновой подложке и начиналось выселение клеток по периферии эксплантата, составляющих зону роста от края эксплантата. Как известно, развитие зоны роста культивируемых фрагментов тканей зависит от пролиферативных, миграционных и адгезивных свойств различных типов клеток. В данной работе учитывали суммарный эффект этих процессов, выражающийся в размерах развивающейся зоны роста эксплантатов. Через 3 сут культивирования в каждой серии экспериментов зона роста характерно изменялась под влиянием какого-либо из пептидов, что давало возможность судить о направленности действия исследуемых веществ.
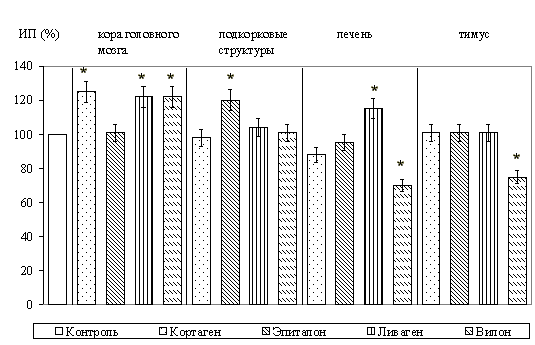
Животные в возрасте 1 сут. Результаты исследований представлены на рисунках 1 и 2.
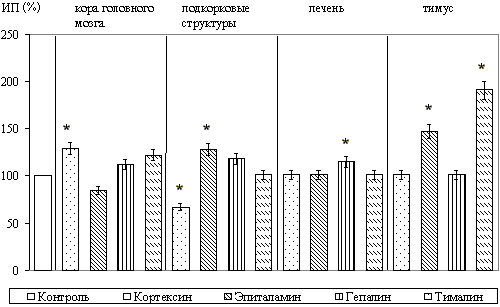
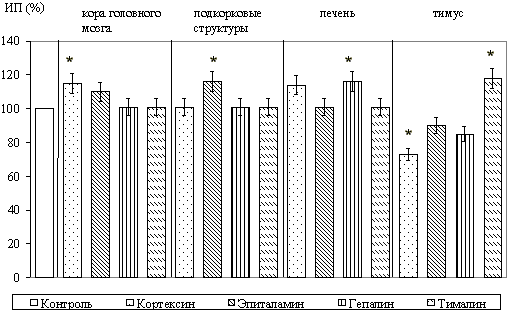
При культивировании фрагментов коры головного мозга в зоне роста отмечалось появление мигрирующих нейронов с крупными центрально расположенными ядрами, а также глиальных клеток и фибробластоподобных элементов. Установлено, что кортексин стимулировал рост эксплантатов коры головного мозга. При этом, ИП повышался на 29,1+/-3,3% (P<0,05) по сравнению с контролем. Из синтетических пептидов рост эксплантатов коры головного мозга крыс стимулировал только кортаген. ИП при культивировании с этим пептидом повышался на 38,5+/-7,1% (P<0,05) по сравнению с контролем. Вилон на рост эксплантатов оказывал достоверное ингибирующее действие.
При культивировании фрагментов подкорковых структур головного мозга, в которых зона роста состояла из клеточных элементов, аналогичных выявляемым в зоне роста эксплантатов коры, выраженная стимуляция роста эксплантатов происходила под влиянием эпиталамина; ИП возрастал на 28,5+/-5,1% (P<0,05) по сравнению с ИП контрольных эксплантатов. Культивирование с кортексином оказывало ингибирующее действие на рост эксплантатов. При исследовании действия синтетических пептидов в культуре подкорковых структур стимуляцию роста вызывал только эпиталон; ИП повышался на 36,7+/-5,1% (P<0,05) по сравнению с контролем. Вилон вызывал торможение роста эксплантатов.
При культивировании фрагментов печени, у которых в формировании зоны роста участвовали гепатоциты и фибробласты, выявлено, что только гепалин приводил к достоверному увеличению ИП эксплантатов на 15,9+/-3,1% (P<0,05). Из синтетических пептидов только ливаген значительно стимулировал зону роста эксплантатов; ИП при его применении повышался на 24,5+/-3,1% (P<0,05) по сравнению с контролем.
При культивировании ткани тимуса зона роста образовывалась за счет мигрирующих и пролиферирующих лимфобластов и некоторого количества фибробластов. Выявлено, что тималин оказывал наиболее выраженное стимулирующее действие на рост эксплантатов тимуса. ИП при этом увеличивался на 91,3+/-5,1% (P<0,05) по сравнению с контролем. Другие комплексы пептидов не оказывали существенного влияния на развитие органотипической культуры вилочковой железы, за исключением эпиталамина, который также стимулировал рост эксплантатов (ИП повышался на 47,3+/-10,1%, P<0,05). При действии вилона в культуре ткани тимуса зона роста уменьшалась на 35,7+/-5,1% (P<0,05).
Животные в возрасте 21 сут. Результаты исследований представлены на рисунках 3 и 4.
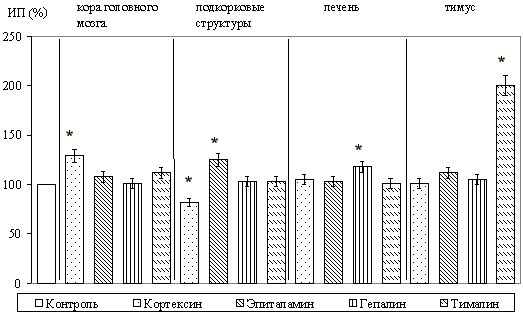
Культивирование с кортексином вызывало достоверную стимуляцию роста эксплантатов: ИП повышался на 27,1+/-5,1% (P<0,05). Из синтетических пептидов на исследуемые эксплантаты стимулирующее действие оказывал только кортаген (ИП повышался на 40,1+/-4,7%, P<0,05).
Выраженная стимуляция роста эксплантатов подкорковых структур головного мозга происходила под влиянием эпиталамина: ИП возрастал на 25,7+/-3,1% (P<0,05). Другие полипептидные препараты не оказывали достоверного влияния на эти эксплантаты, за исключением ингибирующего действия кортексина. При исследовании синтетических пептидов в культуре подкорковых структур стимуляцию роста вызывал только эпиталон (на 27,6+/-3,1%, P<0,05).
При культивировании фрагментов печени выявлено, что только гепалин приводил к достоверному увеличению ИП эксплантатов (на 18,3+/-3,5%, P<0,05). Из исследуемых синтетических пептидов зону роста эксплантатов стимулировал ливаген. При этом ИП повышался на 21,1+/-2,5% (P<0,05).
При культивировании ткани тимуса установлено, что тималин оказывал выраженное стимулирующее действие в культурах тканей; ИП увеличивался на 100,5+/-10,3% (P<0,05). Вилон в культуре ткани вилочковой железы усиливал развитие зоны роста на 15,7+/-0,35% (P<0,05).
Животные в возрасте 24 мес. Результаты исследований представлены на рисунках 5 и 6.
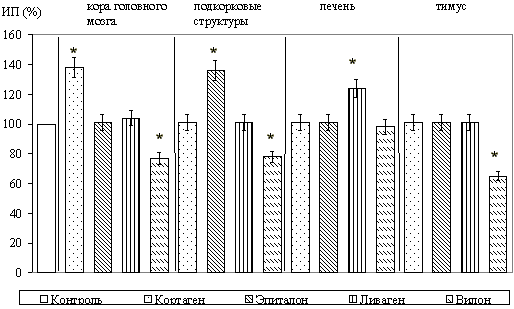
Стимуляция роста эксплантатов коры головного мозга отмечалась только при культивировании с кортексином. ИП повышался при этом на 15,5+/-1,1% (P<0,05). Синтетические пептиды оказывали более сильное воздействие на рост эксплантатов коры головного мозга старых животных, чем это наблюдалось у молодых животных. Это выражалось в том, что не только кортаген вызывал достоверное увеличение ИП (на 25,7+/-4,5%, P<0,05), но аналогичным действием обладали также ливаген и вилон.
В культурах подкорковых структур старых животных стимуляция роста эксплантатов происходила под влиянием эпиталамина; ИП возрастал на 16,9+/-4,5% (P<0,05). Кортексин не оказывал того тормозящего влияния на рост эксплантатов, какое он оказывал на эксплантаты коры головного мозга молодых животных, когда ИП снижался в среднем на 18-33% по сравнению с контролем. Эпиталон вызывал достоверное увеличение ИП эксплантатов подкорковых структур старых крыс на 20,1+/-2,7% (P<0,05). Однако ингибирующее воздействие вилона на подкорковые структуры головного мозга, которое проявлялось в исследовании на эксплантатах животных в возрасте 1 сут, не отмечалось.
При культивировании фрагментов печени старых животных обнаружено, что гепалин приводил к достоверному увеличению ИП эксплантатов на 15,7+/-1,1% (P<0,05). Ливаген также вызывал стимуляцию роста эксплантатов на 15,5+/-0,7% (P<0,05). Вилон оказывал ингибирующее действие на рост эксплантатов печени у старых животных.
При культивировании ткани вилочковой железы старых крыс тималин оказывал не такое выраженное стимулирующее действие, как у молодых животных. ИП увеличивался на 18,3+/-3,1% (P<0,05). При этом, другие исследуемые полипептиды не оказывали влияния на развитие органотипической культуры вилочковой железы, за исключением угнетающего действия кортексина. Вилон угнетал зону роста эксплантатов тимуса старых крыс на 25,1+/-3,1% (P<0,05).
Таким образом, полученные данные свидетельствуют о том, что выделенные из различных тканей комплексы пептидов и синтетические пептиды оказывают тканеспецифическое действие на рост эксплантатов соответствующих тканей как молодых, так и старых животных. При этом эффекты пептидов имеют выраженную зависимость от используемых концентраций и в ряде случаев различаются, в зависимости от возраста экспериментальных животных. Известно, что процесс старения характеризуется сложным комплексом молекулярно-генетических и биохимических изменений, сопровождающихся дезорганизацией пептидергической системы регуляции функций организма. При изучении процессов возрастной инволюции органов и тканей организма выявлено снижение в них интенсивности синтеза белка, продукции физиологически активных веществ пептидной природы и ослабление чувствительности к ним клеток-мишеней, что позволило сделать вывод о важной роли пептидов в регуляции механизмов старения. Результаты биотестирования пептидов с использованием в качестве экспериментальной модели эксплантатов тканей крыс разного возраста свидетельствуют о том, что с возрастом ткани-мишени становятся чувствительными не только к воздействию пептидов родственного происхождения, но и к некоторым другим. Установленные факты свидетельствуют о координирующем действии пептидов на уровне межклеточных взаимодействий, нарушаемых в процессе старения.
Вместе с тем, установленные тканеспецифические эффекты исследуемых пептидов, несмотря на их гипотетически родственное происхождение, не находят окончательного объяснения. Результаты исследований на тканевом уровне позволяют лишь констатировать феномен тканеспецифичности, не позволяя выявить ее механизмы. Известно, что некоторые биологические эффекты регуляторных пептидов основываются на лиганд-рецепторных взаимодействиях или на непосредственном проникновении пептидов в клетку. В то же время, не находят объяснения механизмы как полифункционального, так и тканеспецифического действия пептидов, а также эффективности их применения в сверхмалых дозах [19, 20]. Можно предположить, что для коротких пептидов, образующихся в организме в результате протеолиза или вводимых извне, характерен механизм дистантной "волновой" передачи семантической информации. В этом случае взаимодействие на основе явления резонанса может объяснить некоторые тканеспецифические эффекты пептидов, исходя из родственной взаимосвязи "пептид-ткань".
Известно, что информационные отношения как внутри отдельных функциональных систем, так и в межсистемных связях в целом организме весьма чувствительны к различным воздействиям [2]. Учитывая тот факт, что старение сопровождается изменением экспрессии тканеспецифических генов [14, 22], следствием которого является нарушение синтеза регуляторных тканеспецифических пептидов в различных органах и тканях, пептиды можно рассматривать в качестве информационных регуляторов генетической стабильности. Следует полагать, что характерным проявлением тканеспецифического действия пептидов является совокупность преобладающих биологических эффектов, проявляющихся на различных уровнях организации соответствующих функциональных систем организма: субклеточном, клеточном и тканевом. В основе этих процессов лежит дифференцированная селекция информационных сигналов пептидных молекул.
Работа выполнена при поддержке РФФИ (грант N 01-04-49529).
Литература
- Аль-Шукри С.Х., Горбачев А.Г., Кузьмин И.В., Хавинсон В.Х. Введение в биорегулирующую терапию при урологических болезнях. - СПб.: Наука, 1996. - 93 с.
- Зилов В.Г., Судаков К.В., Эпштейн О.И. Элементы информационной биологии и медицины. - М.: МГУЛ, 2000. - 248 с.
- Кузник Б.И., Морозов В.Г., Хавинсон В.Х. Цитомедины: 25-летний опыт экспериментальных и клинических исследований. - СПб.: Наука, 1998. - 310 с.
- Морозов В.Г., Хавинсон В.Х. Новый класс биологических регуляторов многоклеточных систем - цитомедины // Успехи соврем. биологии. - 1983. - Т. 96, вып. 3. - С. 339-352.
- Морозов В.Г., Хавинсон В.Х. Роль клеточных медиаторов (цитомединов) в регуляции генетической активности // Изв. АН СССР. Сер. биол. - 1985. - N 4. - С. 581-587.
- Морозов В.Г., Хавинсон В.Х., Малинин В.В. Пептидные тимомиметики. - СПб.: Наука, 2000. - 158 с.
- Рыжак Г.А., Малинин В.В., Платонова Т.Н. Кортексин и регуляция функций головного мозга. - СПб.: Фолиант, 2001. - 160 с.
- Хавинсон В.Х. Способ получения пептидов, обладающих тканеспецифической активностью, и фармацевтические композиции на их основе. - Патент РФ N 2161501. - 2001.
- Хавинсон В.Х. Тканеспецифическое действие пептидов // Бюл. эксперим. биол. и медицины. - 2001. - Т. 132, N 8. - С. 228-229.
- Хавинсон В.Х., Морозов В.Г. Пептиды эпифиза и тимуса в регуляции старения. - СПб.: Фолиант, 2001. - 160 с.
- Хавинсон В.Х., Морозов В.Г., Чалисова Н.И., Окулов В.Б. Влияние пептидов головного мозга на клетки нервной ткани in vitro // Цитология. - 1997. - Т.39, N 7. - С. 571-575.
- Хавинсон В.Х., Хокканен В.М., Трофимова С.В. Пептидные биорегуляторы в лечении диабетической ретинопатии. - СПб.: Фолиант, 1999. - 120 с.
- Хавинсон В.Х., Чалисова Н.И., Морозов В.Г., Малинин В.В. Роль пептидов тимуса в регуляции роста лимфоидной ткани у крыс разного возраста // Докл. АН. - 1999. - Т. 369, N 5. - С. 701-703.
- Barazzoni R., Nair K.S. Changes in uncoupling protein-2 and -3 expression in aging rat skeletal muscle, liver, and heart // Am. J. Physiol. Endocrinol. Metab. - 2001. - Vol. 280, N 3. - P. E413-419.
- Diaz D., Krejsa C.M., White C.C. et al. Tissue specific changes in the expression of glutamate-cysteine ligase mRNAs in mice exposed to methylmercury // Toxicol. Lett. - 2001. - Vol. 122, N 2. - P. 119-129.
- He B., Deckelbaum R.A., Miao D. et al. Tissue-specific targeting of the pthrp gene: the generation of mice with floxed alleles // Endocrinology. - 2001. - Vol. 142, N 5. - P. 2070-2077.
- Ivanov V.T., Karelin A.A., Philippova M.M. et al. Hemoglobin as a source of endogenous bioactive peptides: the concept of tissue-specific peptide pool // Biopolymers. - 1997. - Vol. 43, N 2. - P. 171-188.
- Karelin A.A., Blishchenko E.Yu., Ivanov V.T. A novel system of peptidergic regulation // FEBS Lett. - 1998. - Vol. 428, N 1-2. - P. 7-12.
- Khavinson V.Kh., Goncharova N., Lapin B. Synthetic tetrapeptide epitalon restores distributed neuroendocrine regulation in senescent monkeys // Neuroendocrinology Lett. - 2001. - Vol. 22. - P. 251-254.
- Khavinson V.Kh., Izmaylov D.M., Obukhova L.K., Malinin V.V. Effect of epitalon on the lifespan increase in Drosophila melanogaster // Mech. Ageing Dev. - 2000. - Vol. 120. - P. 141-149.
- Morozov V.G., Khavinson V.Kh. Natural and synthetic thymic peptides as therapeutics for immune dysfunction // Int. J. Immunopharmac. - 1997. - Vol.19, N 9/10. - P. 501-505.
- Upadhyay R., Gupta S., Kanungo M.S. Trans-acting factors that interact with the proximal promoter sequences of ovalbumin gene are tissue-specific and age-related // Mol. Cell Biochem. - 1999. - Vol. 201, N 1-2. - P. 65-72.
- Varda-Bloom N., Shaish A., Gonen A. et al. Tissue-specific gene therapy directed to tumor angiogenesis // Gene Ther. - 2001. - Vol. 8, N 11. - P. 819-827.
- Weindruch R., Kayo T., Lee C.K., Prolla T.A. Microarray profiling of gene expression in aging and its alteration by caloric restriction in mice // J. Nutr. - 2001. - Vol. 131, N 3. - P. 918S-923S.
- Xu X.C., Howard T., Mohanakumar T. Tissue-specific peptides influence human t cell repertoire to porcine xenoantigens 1 // Transplantation. - 2001. - Vol. 72, N 7. - P. 1205-1212.
Рис. 1. Влияние кортексина, эпиталамина, гепалина и тималина на рост эксплантатов крыс в возрасте 1 сут. (7,65кб, 500x307 GIF)
* - P<0,05 по сравнению с контролем.
Рис. 2. Влияние кортагена, эпиталона, ливагена и вилона на рост эксплантатов крыс в возрасте 1 сут. (8,96кб, 514x309 GIF)
* - P<0,05 по сравнению с контролем.
Рис. 3. Влияние кортексина, эпиталамина, гепалина и тималина на рост эксплантатов крыс в возрасте 21 сут (7,88кб, 523x316 GIF).
* - P<0,05 по сравнению с контролем.
Рис. 4. Влияние кортагена, эпиталона, ливагена и вилона на рост эксплантатов крыс в возрасте 21 сут. (9,50кб, 517x313 GIF)
* - P<0,05 по сравнению с контролем.
Рис. 5. Влияние кортексина, эпиталамина, гепалина и тималина на рост эксплантатов крыс в возрасте 24 мес. (9,64кб, 512x316 GIF)
* - P<0,05 по сравнению с контролем.
Рис. 6. Влияние кортагена, эпиталона, ливагена и вилона на рост эксплантатов крыс в возрасте 24 мес. (9,88кб, 535x345 GIF)
* - P<0,05 по сравнению с контролем.
V.Kh. Khavinson, V.V. Malinin, N.I. Chalisova, E.I. Grigoriev
TISSUE-SPECIFIC EFFECT OF PEPTIDES IN THE TISSUE CULTURE OF RATS OF VARIOUS AGE
St. Petersburg Institute of Bioregulation and Gerontology of the North-Western Branch of the Russian Academy of Medical Sciences
3, Dynamo Prospect, 197110 St. Petersburg, Russia, e-mail: vvm@medport.ru
I.P. Pavlov Institute of Physiology of the Russian Academy of Sciences
6, Makarov Embankment, 199034 St. Petersburg, Russia
There was studied the effect of polypeptide preparations isolated from the cerebral cortex (Cortexin®), epiphysis (Epithalamin®), liver (Hepalin®), thymus (Thymalin®) and small synthetic peptides - Cortagen®, Epitalon®, Livagen®, Vilon® on the development of explants from the cerebral cortex, brain subcortical structures, liver and thymus of rats of various age. With respect to the applied concentrations of the investigated peptides, they exerted tissue-specific effects in the organ-typical culture of animal tissues.
Страница 278 
|

 Articles
Articles
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
 Свидетельство о регистрации сетевого электронного научного издания N 077 от 29.11.2006
Свидетельство о регистрации сетевого электронного научного издания N 077 от 29.11.2006
